

Livebearer intro
Evolution of viviparity
The genome sequence now decoded of one of the ovoviviparous teleosts, the platyfish "Xiphophorus maculatus", provides a unique insight into the evolution of inner gestation (Schartl et al., 2013). It has been considered that evolutionary transitions occur from ovuliparity and oviparity evolved to viviparity (Lodé, 2012). Interestingly, 13 homologs of mammalian genes related to placental development were found in platyfish, and three of them seemed to be under positive selection. These results suggest that the mammalian and teleostean placentas are convergent, not homologous, organs (Schartl et al., 2013)..
Are fish coming from tropical countries always tropical fish or also subtropical? Fish species (so, also livebearers) occur in a range of fish families and are to be found in multiple parts of the world. All have their own requirements and behaviors. Most known livebearers are endemic to South- and Central-America and Southeast Asia. But because of the fact that endemic locations seem tropical to a lot of people, livebearers are considered being tropical fish. Well, that's an incorrect statement to make! Yes, there are tropical livebearers but there are also subtropical livebearers. And even within the tropical zone on this globe there are areas that have a subtropical climate. But there are also livebearers living in cold water environments. Specifically, certain altitudes won't have the tropical conditions in a tropical global zone. Most livebearers found elsewhere on this globe are mostly feral strains..
Most people think that livebearers have optimal conditions in hard water. Well, this is not a correct statement. Yes, most livebearers in the wild live in hard water conditions but there's also a fair number of livebearer species that live in soft water conditions. Even a decent number of wellknown common aquarium livebearer species can be adjusted to soft water conditions. So, don't say that all livebearers need hard water to live in. For that would be a wrong statement to make. In order to make such a statement, one need to know with what kind of livebearer we're dealing with..
Easy beginners fish...?
A remark that's made so often by people, is to claim that livebearers are the easiest fish to keep and that they are like rabbits when it comes to reproduction (referring to speed of reproduction and number of fry). Well, as a serious livebearer keeper and breeder and referring to all livebearer species there are, I do need to contradict that. For the remark that they are so easy within that field, only relates to a certain number of them (and to most people that remark relates to the known commercial livebearer species). For there are a serious number of livebearer species that are less reproductive and harder to maintain. So, claiming that all livebearers are easy to maintain and to breed should not be seen as a correct statement. And the remark that livebearers are real beginner's fish is only suited for the commercial known livebearers. As already stated before, there's a serious number of livebearers that are harder to maintain. Someone who's claiming differently is clearly lacking knowledge..
By most people most known livebearers are guppies, mollies, swordtails and platies which belong to the Poeciliidae family. However, there are a lot more livebearer species (poeciliid fish and others) which are kept by aquarists and found in nature. The majority of them are considered being rare livebearers. And most are kept by serious and experienced aquarists.
Most people do keep the cultivated domestic species of the Poeciliidae family at home. And specifically from the genus Poecilia." Poecilia" is derived from the greek word "poikilos' which means lots of colors. But I can assure you that not all members of the wild populations within the Poecilia genus are colorful.
Phenotypes
I also would like to refer to phenotypes within a wild population. If we look at oviparous fish (egglayers) of a certain location in the wild, "most" populations will consist of specimens with the same phenotype. Same body shape, same body color, same pattern and same fin shape. But when we look at ovoviviparous and viviparous livebearer popualtions in the wild, "most" of these species can consist of multiple phenotypes.depending on the livebearing species, they can differ in fin shape, body color, body pattern and what I've notice by own experience even some in body shape (which has to do with external factors).

Above: A sample of phenotypes which can occur in one and the same population of Skiffia multipunctata.
Below: A sample of different phenotypes of Xiphophorus hellerii.

As you can see in the sample above, not all Xiphophorus helleriis are real green specimens despite of the fact that we call the Xiphophorus hellerii the green swordtail. These are just some of the phenotypes that occur within the wild Xiphophorus hellerii. So, think again whether the Xiphophorus hellerii should be labeled as the "green swordtail". Look at the sample below of a green Xiphophorus hellerii.
The ones shown above are from different localities. But also with this swordtail species "Xiphophorus hellerii" goes that within an individual population multiple phenotypes "can" occur.

Livebearers come in several sizes...
As already stated, livebearers come in several sizes. A number of sharks or stingrays are livebearers as well. But aren't toothcarps. But they can become very large. There are also livebearers that belong to the smallest livebearing toothcarps. These are: Heterandria formosa (often incorrectly labeled as the smallest livebearer), Neoheterandria elegans, Phallichtys tico, Quintana atrizona , Poecilia scalpridens and Pamphorichthys hollandi (also known as Pamphorichthys minor) . These are all similar in size.
It's been stated that there are diffences in size of these dwarf livebearers. So, there must be a distinct smallest livebearer species. But in reality it has turned out that males differ in adult size and so do the females. Whether that's because of the food they get, at which temperature they grew up or another factor, reality will show that no matter which of these dwarf livebearers it may concern, there will always be differences in adult size in each dwarl livebearer species (no matter male or female). So, I wouldn't prefer to refer of the smallest livebearer species but about "one" of the smallest livebearer species instead.

Above: Phylogenetic tree of 94 species of the Poeciliidae family.
The arrow on the scale bar corresponds to a matrotrophy index value of 1.0, which indicates the division between lecithotrophic and placentotrophic species.
The current analysis suggests that the common ancestor of the family has a placenta and that there were multiple losses and gains of placentation within the family." (red: no placenta; green: poorly developed placenta; blue: well-developed placenta; Figure taken from Pollux et al., (2014)).
I also like to use the words "wild" and "fancy" livebearers. Within the common trade, names like fancy, commercial, domestic and even hybrid are used for the same kind of livebearer. So, what's the difference? Commercial would refer to a product for the market, hybrid would imply a hybridization between two different species, domestic is a specific term used for organisms bred with a temperament useful for humans. To get away from these distinctions, I prefer to use the term "fancy" instead for all breeding forms that are derived from a wild livebearer species.

A serious number of wild livebearers are also introduced in other waters around this globe. Mainly for a specific purpose like combating certain natural pests. Some good examples are guppies and mosquitofish. But also other livebearer species are used for this purpose. But several wild livebearer species were also introduced in other waters by man just as a new species. But the most important condition to do this is that such livebearers should be easy to adapt to other circumstances (e.g., water parameters like temperature and such...). For not all livebearer species are that adaptable.
There are also freshwater systems that are thermally influenced and provide suitable conditions for non-native species of tropical and subtropical origin to survive and form proliferating populations beyond their native ranges.
Below: A monument constructed in Sochi (Russia), honouring the western mosquito fish for eradicating malaria in the region.

In general fish are wether oviparous or viviparous. When fish are oviparous, the females will spawn the eggs. The fertilization can happen internal (true oviparity) or external (ovuliparity). The ability of a female to produce egg cells is called "oogenesis". Oogenesis happens without any stimulation of a mating or sperm.
When it comes to livebearers, they won't lay eggs (oviparous) but have embryonic development within the mother after an internal fertilization. And when the female is due, she'll give birth to free swimming offspring.
To most people, livebearers are all alike when it comes to reproduction. But that ain't correct for there are two ways how these kinds of fish reproduce. Most known are the ovoviviparous livebearers like e.g., mollies, guppies, swordtails, platies and so on... Such livebearers are also called "pseudo livebearers". But there are also the true livebearers like e.g., goodeids (splitfins), halfbeaks, etc. These true livebearers do have a reproduction which is pretty similar to mammals. Livebearers like these are called viviparous livebearers.
True viviparous livebearers like goodeids in the embryonic state are being nourished by the mother through the socalled trophotaenia (or also spelled as trophotaeniae). It's in some way similar to the umbilical cord (placental viviparity) in mammals. These trophotaenia will be shed or absorbed (they'll nourish themselves through the trophotaenia) in the first till two days after birth. These trophotaenia do look like a cluster of small umbilical cords.


Above: A heavily pregnant female goodeid and almost ready to drop her fry.


Above: An illustration of the inside of a pregnant goodeid (Goodea luitpoldi). The space is used optimal by the fry.
Left: A fry of of the Xenotoca eisini.

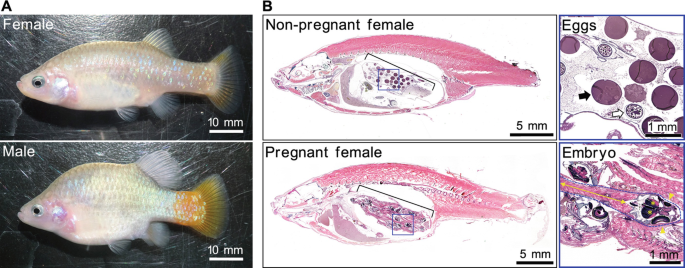
With ovoviviparous livebearers, the embryos have no placental attachment and will be totally nourished from the yolk sac. So, the mother does not nourish the embryos, she's just a safe place before the eggs will hatch. By the time that the mother is ready to give birth, the eggs will hatch. and the fry will pop out of the female's body.
Mostly they're curled up when they leave the mother's body. But sometimes when it takes a bit longer before fry can leave the mother's body, the rear end of the fry will come out first.
The fry do have a big belly (this is called the yolk sac). They'll nourish themselves the first till two days after birth with that what they've stored in their yolk sac. And then they'll slim down and then they'll look for food themselves.
A lot of livebearers are known to reproduce well in captivity. There are exceptions to this rule and some livebearers are very hard to breed in captivity. They might require particular circumstances in order to get them to breed. If the right conditions are not given to them, some female livebearing species will often miscarry, give birth to the embryos too soon or even absorb the embryos.
Since the male livebearer has to fertilize the eggs internally, all male livebearers have an anal fin and this has developed into a reproductive organ that can be inserted into the female. With males of an ovoviviparous species, the reproductive organ is called "gonopodium". Males of a viviparous species e.g., from the families "Goodeidae" and "Hemirhamphidae", the reproductive organ is called "andropodium". Note that the andropodium is just a sort of notch on the front part of the anal fin.

One of the main differences between the two male sexual organs is that an andropodium wil fold when the male inserts it into the female, while a gonopodium will flip forward to insert the female as shown in the picture below. But it's not a full penetration. Actually, the male only connects at the front of the vent (urogenital pore of the female) where he deposits his sperm in just a second. The sperm will be sucked in by the female herself where she will store the sperm randomly as small packets in the folds of her fallopian tube. While the female herself will decide when she will release one or more sperm packets to fertilize her eggs, the use of sperm packets wil be used randomly. So, it's not that the first donated sperm packets or the last donated sperm packets will be used first. So, don't rely on it when someone claims that the first or the last donation will be used first. It really happens randomly. If a female has mated with more than one male, the outcome of one batch of future fry can be fathered by multiple males instead of just one. But as long as a female has not used one or more sperm packets, she won't be pregnant. For I do read and hear so often that livebearer females are always pregnant, which isn't true. A lot of them do get easy pregnant, but that's something different than being pregnant all the time.
Note: An ovoviviparous livebearer female can store sperm packets for over a year without any loss of the quality of the sperm. So, remarks like an ovoviviparous female livebearer can store sperm packets for only a couple of months is totally incorrect..! She won't be clean (no storage of sperm) after a couple of months...! Please, keep that in mind...!


Above: Examples of male mating behavior in different poeciliid species. Courtship in (A) Girardinus metallicus and (B) Limia nigrofasciata. (C) A male Heterandria formosa approaches a female from behind, in this non-courting species. Mating attempts in the form of gonopodial thrusts in (D) Poecilia parae, (E) Phallichthys quadripuctatus, and (F) Poeciliopsis occidentalis.
Photo credits: (A) https://acquariofiliaconsapevole.info/pesci_girardinus_metallicus, (B) Katie Holz, (C) TFH Archives, (D) Godfrey Bourne, www.discoverlife.org, (E) http://aquaria.palo-alto.ca.us/, (F) Dennis Caldwell.
Many people think that keeping multiple kinds of livebearers in one tank will create inbreeding. Most of the time that will not be the case. First of all, when it comes to ovoviviparous livebearers, the shape of the gonopodium of each species does differ.
And the female sexual organ is adjusted to the gonopodium of a male of her own kind. In order to interbreed with another livebearing species, the sexual organs need to be compatible. But if two species are closely related to eachother, the bigger the chance the sexual organs will be compatible.
Are there both genders of two or more different livebearing species in one tank, their first preference is always to mate with one of their own kind if both genders of that particular species are present. Are there just opposite genders of a specific species present, only than it's most likely that a crossbreed with a sexual compatible tankmate of the opposite sex can be established.

The opening of the sexual organs of both genders are called gonopores.
The difference between the global shape of the anal fin between male and female of both ovoviviparous livebearers will be shown by the pictures below.


Looking at these pictures shown above, it's obvious that there is a clear difference in the shape of the anal fin of both genders. And also wether it concerns an ovoviviparous livebearer or a viviparous livebearer. So again, don't think that all male livebearer specimens have a gonopodium. And that not all female livebearer specimens have a V-shaped anal fin. For that would be a complete misunderstanding!
Resumé:
The structure of both male and female sex organs differ in structure in livebearer species. Only if both male and fremale sexual organs are compatible, an actual mating can take place. This disables "a lot of livebearer species" to mate with another. So, it's not always the genera they're in that makes the difference, also the compatibility in sexual organs.is of importance.
Early sexing of the genders is possible for a number of ovoviviparous livebearers. For viviparous livebearers it's harder to determine the correct sex at an early stage. Ovoviviparous livebearers that can be sexed at an early age are to be recognized by the shape of the anal fin. The first finrays of a male is slightly bend and of a female the first finrays are more straight. A bit later the tip of the anal fin of a very young male is a bit pointy and a bit round when it concerns a female. But you need to have a good eye for it to recognize it. Also a large number of very young ovoviviparous female livebearers will show a tiny gravid spot already after a couple of days of their lives. But be aware that not all ovoviviparous female livebearers will show a gravid spot (not even when they're pregnant!). But the majority of those ovoviviparous female livebearers will show one.

Above: A week old female fry that shows the gravid spot already.
And to refer to the different shapes of the gonopodium again (a bit earlier I´ve used explicitly "global shape" for even the shape of the gonopodium of almost each ovoviviparous livebearer male differs in shape)... Here are some pictures which show how different those gonopodiums are shaped depending on the livebearer species.


If you'd enlarge these pictures, you'll be able to see clearly that gonopodiums do differ in shape from the start to the tip of the gonopodium. This is very essential to the gonopore of the female to enable a mating. This is one of the main reasons why not all ovoviviparous livebearers can crossbreed with another.
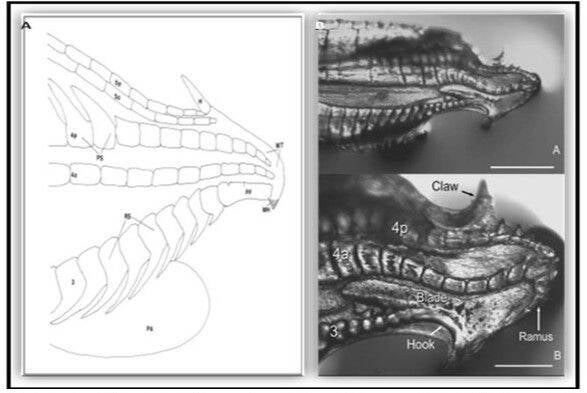
Above & below: Gonopodium structure observation of Poeciliidae.



Above: The tip of the male gonopodium, the intromittent organ, exhibits a great deal of variation across the family Poecillidae: (A) Poecilia reticulata, (B) Xiphophorus helleri, (C) Phalloceros caudimaculatus, (D) Belonesox belizanus, (E) Girardinus cubensis, (F) Neoheterandria elegans, (G) Heterandria formosa, and (H) Poeciliopsis elongata.
Relative gonopodium length in the family Poeciliidae ranges from 14 to 45% of male standard length. (A) Males of Poecilia wingei and Limia perugiae (pictured left) have relatively short gonopodia while males of Phalloptychus Januarius and Poeciliopsis elongata (pictured right) have relatively long gonopodia. (B) Frequency histogram of male gonopodium length (n = 92). (C) Ancestral state reconstruction of relative gonopodium length as a continuous character using the contMap function in the R package phytools, which estimates the Maximum likelihood ancestral states at all internal nodes and interpolates the states along each edge. The phylogeny tips depict a binary categorization of relative gonopodium length (i.e., short vs. long). The most likely ancestral state of the family, when treated as a binary character, is that of a long gonopodium (Furness et al., 2019).

Ovoviviparous male livebearers with a long gonopodium won't have a real courtship when trying to mate. For they can oversee the mating attempt. The end of their gonopodium is also more smoother shaped than a gonopodium of a male with a shorter gonopodium. Ovoviviparous male livebearers with a shorter gonopodium do have a courtship to persuade the female to mate and so they can come closer to the female's gonopore. Because it needs more work for a male with a shorter gonopodium, the end of his gonopodium has got 1 or more hooks to have a better copulation. This can be exhausting for some male specimens. Sometimes it can even happen that they get stuck in the gonopore of the female. Which can lead to stress to both male and female. Worse scenario is that they can get injured and it can even be traumatic.
Note: In general, when an anal fin of a young male will develop into a gonopodium, the anal fin starts to change from the third, fourth and the fifth fin ray. But there are also specimens that start the transition by thickening the first anal finrays.
Below: The anal fin starts already to be pointy at the tip. This young male platy is developing a gonopodium.

What also can happen is that an initial ovoviviparous male won't develop a real gonopodium. Such males grow up large as well (like late males do). At some point, the first anal finrays are becoming pointy and longer. But that's all that happens. It's a phenomenon that can happen with every kind of ovoviviparous livebearer. Such males can not breed with a female.

Above & below: An adult large male with an undeveloped gonopodium. With most of such guppies, there's hardly coloration or pattern present. But there are also specimens that do carry color but they'll be paler than their male siblings. With this male shown, you'll also notice that this ain't a female. For a gravid spot was never developed (just like late males).
So, if you 'll see a similar male, don't mix it up as being a female, despite of the female bodyshape.

Below: Also this male Japan blue grass guppy (with zebrinus markings) is not that intensively colored as his siblings. And with 6 months of age, he hasn't developed a proper gonopodium. As you can see, there are no hooks on the tip of the gonopodium as he should have. And only the first finrays are thickened and the finrays following haven't changed. So, also this male can not be used for breeding.


There are also ornamental ovoviviparous livebearers where males have an extreme long gonopodium. Most known ones are in guppies and swordtails. When it comes to guppies, such an extreme elongated gonopodium occurs in socalled ribbon guppies. Ribbon guppies also have two elongated ventral or pelvic fins. The problem with such a fullgrown male is that it's impossible to direct his gonopodium to the female's vent to donate sperm. They're not infertile but they are sexual disfunctional. Only the beginning of the gonopodium (close to the body) can be controlled but not the tip of the gonopodium. Therefore, such a male can not be used for a breeding project.
The same thing goes for swordtails. Lyretail swordtail males can (so, not all lyretails) also develop an extreme elongated gonopodium. Also these kind of males are useless for breeding. See photo below.


But wether it's about lyretail swordtails or ribbon guppies, when you breed those strains, there will be born two kinds of males. Males that will develop an elongated gonopodium and males that will develop a normal shaped gonopodium. The males with a normal shaped gonopodium can be used to breed with a lyretail female. In the next male offspring there will be both males with elongated gonopodiums and normal sized gonopodiums. The males with the elongated gonopodiums are used for shows and just being pretty during their lifetime. Nothing more, nothing less.
But also female ribbon guppies have an elongated anal fin. Female lyretail swordtails however can (so, not always) have an elongated anal fin. But that won't be of any issue when it comes to breeding.

Above: Female ribbon guppy
Right photo: Female lyretail swordtail

Below: Male siblings with normal and elongated gonopodiums.

The gravid spot:
Well, most female livebearers do have a gravid spot. But, there are also female livebearers that won't show a gravid spot. That's why we can state that a gravid spot is not crucial to determine a female livebearer from a male livebearer. The gravid spot is the most translucent part of the skin, close to he anal fin and where the sexual organ of the female is situated. Most females do show this spot already a couple of days after birth. It just needs a good eye to spot this gravid spot at such an early stage. Of course, with female livebearers which are very dark based like e.g., black mollies, even with a very good eye sight, you probably won't be able to spot the gravid spot.
Note: I've mentioned that most female livebearers have a gravid spot for there are also species of which the females don't have a gravid spot.
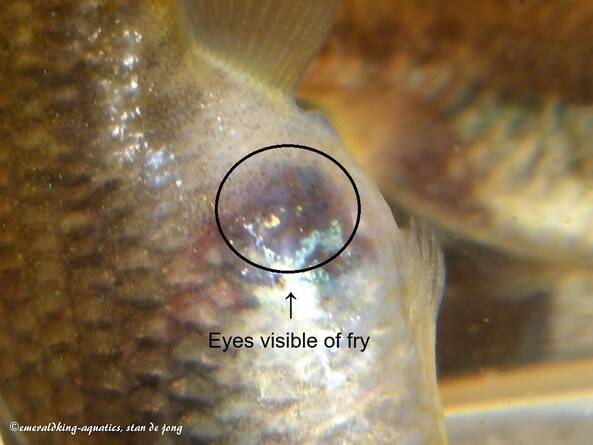
It's also a misunderstanding that when a female's got a gravid spot, that she must be pregnant. It's a misunderstanding that can be read at a lot of forums. Having a gravid spot does not mean pregnancy by all means. For actually, females are born with it. It takes a bit before they show it clearly to repeat myself. As already mentioned, the gravid spot is the most translucent part of the skin. But when the embryonic state develops with a pregnant female, the skin will stretch more and more and so this part of the skin becomes more translucent. What we call the dark gravid spot is basically nothing more than developing embryos that will shine through the translucent gravid spot. And that's why it seems to get darker. At a certain moment you'll be able to see those eyes of the unborn fry through the female's skin.

The coloration of the gravid spot differs per female. Firstly the gravid spot color of a non-pregnant female depends on the base color of the individual female. For instance, grey bodied guppies will have a dark gravid spot, blonde guppy females have a lighter colored gravid spot. But once a female is pregnant, the color of the gravid spot may change. In general grey bodied females, will remain having a dark gravid spot. Blondes, white and golden bodied females, can have a dark or light colored gravid spot when the pregnancy continues to develop. This depends on te fact what kind of base color the bodies of the embryos will be. For instance, if you cross a grey bodied male guppy to a blonde bodied female, the gravid spot will turn dark. For grey is dominant in comparison to blonde. And the fry become grey bodied. Will the embryos be blonde bodied, the gravid spot will be light colored. If the female carries both grey and blonde bodied embryos (let's say that the grey bodied father carries the blonde gene as well), the offspring can consist of grey and blonde bodied fry. Then you'll be able to see a gravid spot on a blonde female that has darker and lighter spots. But a gravid spot can temporarily also change color by emotion, like stress for instance.
Female lacking a gravid spot:
Like already been mentioned by me, there are also females that won't show a gravid spot. What's going on when a female doesn't show a gravid spot? It means that at the spot where usually the gravid spot appears, there is a dense concentration of chromatophores (color cells) in all three skin layers. Which makes that spot not translucent. So, no gravid spot will be seen. A progressing pregnancy won't change anything.
If there are chromatophores (color cells) in one or two layers instead of all three, when the pregnancy progresses, a slight gravid spot can be noticed. But once such a female is done giving birth, you won't be able to see a gravid spot anymore.


Pseudo gravid spot:
Another phenomenon that may occur with xiphophorus strains (mostly platies), is that a juvenile male is developing a socalled "pseudo gravid spot". In general the pseudo gravid spot shows up during the transformation of the anal fin into a gonopodium or after the gonopodium has been formed. The pseudo gravid spot develops in the upper layer of the skin (there are 3 layers) which is totally different than the gravid spot of a female. A pseudo gravid spot hardly shows up before the transition of the anal fin. This has got nothing to do with a female turning into a male. For that's a complete other story.
But there are also other ovoviviparous livebearer species that develop a pseudo gravid spot in males.
Note: When a pseudo gravid spot shows up on a male's body, it will never dissapear.

Above: An adult male Xiphophorus evelynae with a pseudo gravid spot.
Sex ratio:
Another misunderstanding is that you'll need a male:female ratio of 1:2 or 1:3 at all times. The socalled harassment behavior of males goes for just a certain number of livebearers. And most of the times, you see this happen with a certain number of cultivated domestic species. There are a lot of livebearers which can be kept without a problem at an 1:1 ratio in a tank.
For instance, do all male guppies harass females? No! Definitely not! There are a lot of wild but also fancy guppy strains which are just fine at a ratio 1:1. The problem overhere is that over the years we've been taught a simple rule which we've been generalizing over the years for all guppies (also other livebearers of course). Experienced serious aquarists and breeders do know that this generalization is at hand. Unfortunately, this rule and so many others have been copies of statements that someone made up once just to ensure to put the harassment to a low level and we've been continuing these rules as being true.

Reproduction:
Although, a number of male livebearers tend to chase females all day... that doesn't mean that a mating will take place. Even though, a male is trying to use his sexual organ. For it's the female whch decides if a mating will actually happen. Unless, we're dealing with a very young and naive female.
It's also that when a male will copulate with a female, that his gonopodium is not inserting the female's vent like a lot of people may think. He's only connecting on the outside of the female's vent and donates his sperm packets in a split second at the opening of the vent. The sperm packets will be sucked in the female's vent by the female herself.
If a mating does happen, depending on what kind of ovoviviparous livebearer it concerns, the female is able to store sperm in packets for a certain time period. This means that when a mating has happened that the female does not have to be pregnant. She'll decide if she let's a sperm packet fertilize her eggs. And only if that happens, she'll be pregnant. The time period of sperm storing can take as long as over a year. So, not just a couple of months as some like to claim. The total number of pregnancies by one mating, depends on the number of eggs that a female produces each batch and the amount of released sperm packets used each time (this may vary each time). This all means also that such a female can become pregnant multiple times, from 1 -7 times (mostly less). So, not a strict maximum of 3-4 times as I do read frequently. This means also that you can not claim that a female which has stored sperm packets will be clean after a few months. It's possible but not likely. As already been mentioned, sperm storage can even take over a year from one single donation.
Another misunderstanding is that many people will say that an ovoviviparous female livebearer will deliver fry every 3-4 weeks once she's mated with a male or multiple males. This is a complete incorrect information. A large number of females of an ovoviviparous livebearer species will have a gestation period of 3-4 weeks which is incorrectly translated by many people to giving birth every 3-4 weeks. For it's the female herself which decides when she will release one or more sperm packets to fertilize her eggs. So, the time between two pregnancies can variate completely.
The sperm packets are stored in the folds of the fallopian tube (uterine tube). While the female is having this facility and the used folds are opened randomly, you'll never know which sperm packets she will use to fertilize her eggs. So, it could be of an older mating or a recent mating. In case she's having a huge number of eggs, multiple sperm packets will be used to cover the number of present eggs. This means that the offspring of one batch could be fathered by one or more males. If someone likes to linebreed, this is very important to know. That's why virgin and clean (no sperm packets stored anymore) females are so important to use when linebreeding is at hand.
Last in, first used...!
Always last donated sperm, will not always be used first...!
Note: There have been a number of scientific researches that have resulted in the following data: The last donated sperm packets were used to give birth to offspring. Which holds that they were fathered by one particular male. But also I have made my observations and tests throughout the years. And sometimes this goes like what the researches have experienced but often a female opens up randomly a number (or even just one fold) folds of her fallopian tube. Which means that an offspring of one batch can be fathered by multiple males as well. So, in my opinion, those scientific tests were just a lucky shot that the offspring were from one male. They used more males with a different phenotype that were put with the females one after another. So, this means, a couple of days one male after that this male would be taken out and replaced by another male. If all these different males would be put together at once, we wouldn't be sure which one would be a potential father. But they haven't done these tests for years but only during the breeding set up during a shorter period of time. While I've been doing this for decades already. And I also have to mention that eventhough more males with a different phenotype were in the same breeding tanks, it doesn't have to mean that a female has mated with all these males. For the female decides which male she allows to mate with her despite of the fact that those males were trying to mate with her. Also that could be an explanation why only offspring has been born through one particular male. Thusfar, not one research has taking that in account. So, that;s why I also don't take these researches too serious.
Scientific tests don't always have to be a 100% accurate... That's for sure...
And to get to the answer of a certain query, a scientific approach is not always the right answer or the only way to prove something. You don't have to be an official scientist yourself to find out the correct answer to a certain query. But as being ordinairy people we just rely on scientific research before we believe it. And we believe it even more if it's published in a scientific paper or magazine. Also findings as a conclusion of such a scientific test will be published. But that doesn't mean it's proven...!
But how many non scientists have even contributed to certain queries which scientists have benefited from? That's a serious number, I can tell...
If one considers a moment-by-moment explanation as good research, then I think something must be wrong... But despite of my remark, it remains interesting to read what the findings of such a scientific research holds.
And again, the female is the one which decides whether a sperm packet will be used in order to establish a pregnancy. That's why a female is not pregnant at all (even is she's big because of the present eggs) if she's carrying eggs that are not fertilized yet. So, there are big bellies with developing embryos and there are big bellies with just eggs! And therefore the remark that "female ovoviviparous livebearers are always pregnant" should be considered being a false statement.
It will also take only a couple of days or a couple of weeks after fertilization that an embryo has almost fully developed (the exact period differs per livebearer species) . The rest of the gestation time (depending on the kind of livebearer between 3 - 8 weeks) is needed to let the fully developed embryo grow in the egg. The eggs were already provided by the mother with nutrients through follicular epithelia prior to any fertilization (this is called "pre-fertilization").
Sometimes you're able to watch how a pregnant female that is due that she releases eggs instead of fry. Or even eggs and fry. This means that there wasn't sufficient nutrients in the egg yolk. Because in normal conditions all eggs have sufficient nutrients in the yolk. Which also means that the embryonic development of all embryos is at the same pace. A situation when a female is stressing a lot, you'll be able to see the undeveloped eggs coming out first. For they are easier to push out. Those eggs just didn't have sufficient nutrients in the yolk. So, undeveloped eggs have got nothing to do with stress itself. Those eggs would have never developed as being normal embryos starting from the beginning.
The way fertilized eggs develop until birth in ovoviviparous livebearers is called "lecithotrophy". So, there's no maternal care. The mother is just a safe place for the embryos to develop and to grow. Nothing more...
Below: A series of stages of embryo development of a guppy (In vitro culture of a guppy)




Above: An embryo of approximately 3-4 days after fertilization.
Below: This is how a whole bunch of embryos look like in their eggs in the mother's body.


Above: An approx. 6 day old embryo.

Above: The birth of a guppy fry.
Below: A newborn guppy fry. Still having its yolk sac. This will be absorbed within 1-2 days. After it's used all the nutrition in the yolk sac, it will be independently on the search for food. Therefore, it's of no use to feed newborns right after birth.


Above: A three days old siamese guppy twin.
Below: Top view of a one day old still translucent looking guppy fry.
In general, when a conjoined guppy twin (siamese twin) is born, most of the time one of them will grow up normally and the other remains small. Once the twin is full grown, the smaller twin looks like some kind of a tumor hanging at the belly of the other twin. One should not be worried about the healthy fullgrown one. For the tumorlike twin won't be of any burden to the fullgrown twin.

Above: A juvenile conjoined twin. Most of the time the smaller twin will shrink more when the other twin comes to adulthood.
Photo courtesy of David Veshapidze.
Besides conjoined twins, also conjoined triplest can be born. I've seen some that grew up to adulthood. But despite of that fact, they don't tend to have a long lifespan. Conjoined twins has got more chance to live a normal lifespan than conjoined triplets. But most conjoined triplets won't reach adulthood, to be honest. Unless, one or two of the triiplets become parasitic. That way, at least one will become an adult and have more chance on survival.




Above: Conjoined triplets
Why is it of no use to ask how many fry a fish will produce...?
Simply because that number varies every time, no matter if they're egg layers or livebearing toothcarps. I do understand the question involved. But basically, there's no actual answer to it.
I frequently come across this typical question of people: How many fry will this fish have?
A bigger belly in comparison to a less big belly doesn't always mean more eggs or more fry. The eggs can be less but more fluid is in the belly or the fry can be somewhat bigger but less in number when the belly is quite large. But it could also mean more fry but tiny sized in comparison to a normal sized newborn when it comes to livebearers. Although, egg layers and livebearers have there own size of all eggs, if an egg has got more nutrients (embryos need these nutrients to develop), the egg can be bigger than other eggs of the same batch.
The only thing we can say, is that a fish can produce a lot of fry or not... So asking: Why is it of no use to ask how many fry a fish will produce...? Is actually a superfluous question.
Below:
Three heavily pregnant glass belly female guppies. The top photo shows embryos with black eyes (just like the mother). And the bottom photo shows embryos shining through with red eyes (just like the mother). Btw, glass belly female guppies won't show a gravid spot because also their belly is translucent just like a gravid spot itself (lacking color cells overthere). If they're not pregnant, you'll be able to see their guts shining through their belly skin.





With a lot of viviparous and ovoviviparous livebearers, the females become larger when pregnancy goes forward. At the time that they're due, the belly shape can be either round or boxed (also known as squaring off). A lot of people think that when a female is almost due, that the belly will always square off. But that's not true. When a female is squared off, it means that she has used her maximum gestation period. Which is longer than the average gestation period. So, a female that is almost due can also have a round belly (see photo above). It just differs per individual female and if she will take longer before she'll drop her fry. See photo below.

Overhere some photos of an undeveloped egg which had just insufficient nutrients in the yolk. Just some eyes are visible and a bit more (probably the head). The rest is fluid. This abandoned egg measures 3 mm in diameter. In the first and second picture you'll see a just released fry and an undeveloped egg. This concerns a batch of green swordtails.





Above: The undeveloped swordtail egg measures 3 mm in diameter.
Note: The diameter of each egg can differ, no matter the size of the female of each ovoviviparous female livebearer. Which means that also eggs of each batch of the same female can differ in size.
The sperm storage ability goes only for most ovoviviparous livebearers that are lecithotrophic without superfetation and with superfetation. Viviparous female livebearers however have a placenta and don't have the ability to store sperm packets. With pregnancies, the developing embryos are fed through the mother's body (through the placenta) until the actual birth will take place. This method is called "matrotrophy".
But as I've already mentioned "depending on what kind of ovoviviparous livebearer it concerns", means that not all ovoviviparous female livebearers are able to store sperm. Those ovoviviparous female specimens that can't store sperm and the true viviparous female livebearers, do need a new mating in order to start a new pregnancy. So, a resumé: Not all female livebearers are able to store sperm packets!
There's also another way of reproduction with a certain number of livebearer species. The superfetatious livebearers. Superfetation means multiple pregnancies with embryonic development in different stages. Such pregnant females will drop 1-3 fry every single day or 2-3 days during a period of approximately 10-14 days. It's also a different way of sperm storage. Sperm from one mating is only used for one batch of fry during this labour period of 10-14 days. So, a new mating needs to happen in order to establish a new cycle of pregnancy. In this case the pace of development is only at the same pace for each individual small group of embryos but not at the same pace as another small group of embryos that such a female is still carrying.
What's also remarkable is that placental pregnancies of superfetatious female livebearers (which most superfetatious female livebearers are) is also causing that such females start showing bigger bellies later than lecithotrophic females. The hypothesis is that it's got to do with the pace of the waterflow, so that such females are able to swim better with a higher water current for a longer time period.
Below: An illustration of a superfetatious female

Gynogenesis
A unique reproduction method is gynogenesis. It's a bit similar to parthenogenesis but with parthenogenesis it involves the development of a female sex cell (gamete) without fertilization. But with gynogenesis, insemination of an ovum (egg cell) is needed to activate the development of the egg but without the fusion of sperm and the egg nuclei. Which means that the genetic package of the male will not be passed on and so the gynogenetic embryo develops at the expense of the ovum nucleus only. Because of this reproduction method, the fish are asexual (only females). And while the genetic input of a male is of no longer value, the offspring will naturally be all female and identical clones of the mother.
Because of the fact that such fish are females, they need to use sperm from a male related to their own species. A good example of a livebearer which reproduces itself by gynogenesis is the Poecilia formosa also known as the Amazon molly. Such a female needs a male of another molly species to mate with. According to a published article, also other sperm donors than molly species have been used in laboratories to result in successful hybrids with the Poecilia formosa (Schultz & Kallman, 1968; Schlupp, Parzefall & Schartl, 2001; Schlupp, 2010).
Resumé: Gynogenesis is a relatively rare form of asexual reproduction where sperm is required to trigger embryogenesis, but male genes are not incorporated into the genome of the embryo
Do all livebearers prey and eat their fry?
No, would be the correct answer...!
A generalization that's been going on for years, is that adult fish tend to eat fry. Well, that depends on the kind of fish species (in this case livebearers) we're talking about. There are specific livebearer species that will chase and eat fry. But just know that there are also lots of livebearer species that won't chase nor eat fry.
Above: A female Chapalichthys pardalis giving birth. Most goodeids will leave their newborns alone. Also this goodeid doesn't pay attention to the fry. You can really leave those newborns with the adults. No harm will be done.
Above & below: A female Hamburger swordtail is dropping fry. Two different phenotypes were dropped by this female. She doessn't bother at all to chase or even eat her newborns.
It's also a matter of habituation to keep bigger fish with very small ones. Once the bigger fish are used to very small tankmates, they'll start leaving those fry alone. But most people just don't want to invest their time to let those bigger fish get used to those small ones.
It's also a generalization to state that one should always need enough hiding spots for fry to hide in. If the bigger fish leave the fry alone, there's no need for hiding spots. Only if you're dealing with adults that will chase and eat the fry, hiding spots are preferable.
Sex determination:
The majority of animal species can be specified as being bisexual. The genome is in general responsible for deciding wether an embryo will develop as being male or female. A series of developmental sequences are initiated by the sex determination genes which will result into a male or female phenotype. These developmental sequences are called "sex differentiation and which are controlled by gonadal hormones. Sex determination itself is controlled by genes. Male or female phenotype is controlled by so many mechanisms like e.g., the ratio of X chromosomes to autosomes (any of the numbered or non sex chromosomes of an organism).
Sex chromosomes evolve from autosomes and are typically extremely conserved, not only among species but entire classes of organisms. The best-studied sex chromosome systems are ZW and XY, female and male heterogametic systems respectively.
The study of closely related species of fishes that exhibit different sex chromosome systems would be particularly informative for the investigation of the evolution of mechanisms of sex determination. Among the 26 species of the genus Xiphophorus (swordtails and platyfish) are species with both XY and ZW sex chromosome systems. In this genus, these simple heterogametic systems are both present together with more complex situations that can include multiple loci and chromosomes. Xiphophorus maculatus, for example, one of the most well studied and geographically widespread species in this genus, has three different genetically well-defined sex chromosomes, X, Y and W. Possible female genotypes are XX, XW and YW, and in males XY or YY. Several different models have been proposed to explain sex determination with three sex chromosomes in this species. While in the wild in most populations all these sex chromosome combinations co-exist, laboratory lines have been established which are stable for XY/XX or WY/YY sex determination. WW females can be generated in the laboratory after experimental manipulation and are viable, but such individuals have not been reported in natural populations. In several species, autosomal modifiers, which occur at low frequency in natural populations, have also been reported to act, and may explain instances of atypical sex determination. As another example of sex determination variation in this genus, in Xiphophorus hellerii, a polyfactorial sex-determining system has been reported to be acting. Additionally, it has been suggested that this species has a main genetic system affected by numerous autosomal modifiers, while more recent studies report that some strains of X. hellerii have a XW–YY female heterogametic system. To our knowledge, environmental sex determination has not been reported from observations in nature or from laboratory studies in the genus Xiphophorus.
In general, this genus of fish comprises well known and widely studied models for organismic and molecular evolution. In particular, Xiphophorus species have been used as a model system for studying the evolutionary genetics of hybridization for more than 50 years with a recent resurgence of investigations using modern molecular and genomic data
Does water temperature have any influence on the sex of unborn fry?
Officially, there's no scientific proof of water temperature having influence on the sex of unborn fry. But as many (including myself) breeders have observed is that with ovoviviparous female livebearers, the influence of water temperature is present to influence the sex ratio of unborn fry. As far as we know, with viviparous livebearers this trick hasn't resulted in a clear sex manipulation of unborn fry.
In most ovoviviparous livebearers when kept at higher temperatures as they are normally kept at, pregnant females will drop more offspring than female offspring. And when kept at lower temperatures than usuall, more females will be born. But that doesn't go for all livebearer species. As already stated, no clear results with the same method has been found in viviparous livebearers. But in superfetative livebearers such as e.g. Neoheterandria elegans, Heterandria formosa, Phallichthys tico, and so on it's the other way around. When they're kept at higher temperatures in comparison to what they are usually kept at, they'll produce more female than male offspring. And ate lower temperatures than what they are usually kept at m, more male offspring.
Note: This method is not a 100% garantuee that it works but most of the time it does work nicely. And the increase or decrease of the water temperature should be going on during the whole pregnancy otherwise it's of no use. For every single embyos starts of as being as a female state. And while sex determination happens already in the first couple of days after fertilization (so, the beginning of the embryonic state), it's crucial to regulate the water temperature early.

Above: From stage I it's important to increase or decrease the temperature to influence the sex of an embryo.
Sex change in Xiphophorus species:
Overhere a quotation of a paper of Prof.,Dr.rer.nat.,Dr.h.c. Manfred Schartl that will explain the sex chromosomes that Xiphophorus species have. Platy species have besides the sex chromosomes "X" and "Y" also the sex chromosome "W" unlike other ovoviviparous livebearers. The paper was received on December 19th 2003; received in revised form on February 2004; accepted March 3rd 2004.
Quote of "Mechanisms of development- Sex determination in platy fish and swordtails":

The quote "Another modification of the XY system was firstly ound in Xiphophorus nigrensis. In this species all XY fish are males. There is an autosomal locus with two alleles, A and a, which affects sex determination in XX fish. If the genotype is AA, the fish will be female, and if aa, the XX individual will be male. Aa genotypes are predominantly female, but sometimes (approx. 5% during the researches) such fish develop into males (Kallman, 1984)". It has turned out to be the same in other kinds of swordtails. Which explains the sex change in swordtails. And as far as experience have shown, these transformed females will become complete functional males.
An older swordtail female can show a sword-like tail shape and a gonopodium-like anal fin. This is not a late male or an Aa genotype female. This happens by an hormonal change. This hormonal action triggers an overacting stimulans of the MSX-genes. The MSX-genes are responsible for the morphological change in fin tissue (see further text below about the MSX genes). MSX genes are present in both male and female specimens.
XX female platies will always remain females. WY and WX platy females however can change sex at some point. WY platy females that will transform will become functional males (so, they're able to fertilize a female's eggs for they were born with both male and female gonads but will start off as being female). WX females that will transform into a male but won't be functional males afterwards.
All that has been said above about the sex change refers to wild species (so, no hybrids between swordtail and platy species). Most (so, not all) breeding forms are a result of crossbreeding swordtail x platy species. Also sex changes in breeding forms of platies and swordtail fish occur. But if they're derived from a crossbreeding between platy and swordtail, it will be a question whether the sex change will happen as in pure platies or as in pure swordtails. So, don't be surprised when you'll find breeding forms of swordtails that carry a W sex chromosome.
When does a sex change happen?
A sex change mostly happens when there's a trigger. Triggers can be when a group of Xiphophorus species doesn't contain males, there's a significant lack of males or too much dominance of one or more males is present. In last case, more developing males will give a good resistance in the dominance of the other males.

Secondary sexual characteristics evolve in response to sexual selection. A prominent example for this phenomenon had originally been described by Charles Darwin (1871) in fish of the genus “Xiphophorus”, specifically the swordtail fish. In males, the ventral caudal fin rays are strongly elongated and often ornamented with bright pigmentation, forming a conspicuous extension of the caudal fin which resembles a sword (see illustration below under “A”). Note: Not all wild swordtail species will have males that will develop a sword. .lnterestingly, females of the green swordtail (Xiphophorus hellerii) and several other species preferentially associate with males carrying a longer sword over males with a shorter sword, which is thought to result in a higher mating success of long-sworded males.
Within the Xiphophorus genus, several species won’t develop a sword, specifically the platy fish. Note: In some wild platy species, males can develop a sword (mostly very short). The origin of the sword is debated and relies on a robust phylogeny of the genus, which despite various efforts using morphological, electrophoretic and molecular characters is still debated in some details (Rauchenberger et al, 1990; Basolo 1991; Meyer et al, 1994; Borowsky et al, 1995; Marcus and McCune, 1999). Hybridization experiments between platys and swordtails suggested that several ‘”sword genes” are required to induce a caudal fin extension (Zander and Dzwillo, 1969). Although some of these factors might be present in platy fish, in the absence of further sword gene activity, as it is supplied by the swordtail genome in hybrids, fin elongation does not occur (Zander and Dzwillo, 1969). However, the molecular mechanisms underlying both phylogenetic and ontogenetic sword formation and putative subsequent sword loss so far have remained completely unknown.

Juvenile swordtail males are initially swordless. The onset of sexual maturity marks the initiation of the development of secondary sexual characteristics. A sustained increase of testosterone levels during male sexual maturation has been shown to be sufficient to initiate the molecular pathway toward sword development. Moreover, exogenous addition of testosterone can induce a caudal fin protrusion in female swordtail fish, in juvenile males and some normally swordless platy species (see illustration under “C”). During sword development, the ventral caudal fin rays develop with positive allometric growth by adding additional segments of regular segment length to the distal tip of the fin and develop a characteristic pigmentation pattern. Although the sword is an ornamental fin ray modification, specific to the genus “Xiphophorus”, another male structure derived from the anal fin rays, the gonopodium, is a common functional feature of all ovoviviparous members of the “ Poeciliidae family” (see illustration above: A,B and C). The gonopodium arises during sexual maturation in male fish under the influence of testosterone by growth and differentiation of the anterior anal fin rays and serves as a copulatory organ in these ovoviviparous livebearers (Grobstein, 1942). Nothing is known about the molecular pathways on which androgen hormones act during formation of the sword and the gonopodium.
MSX genes and the closely related DLX genes are expressed during all phases of vertebrate development and mediate patterning and morphogenesis in several tissues, including neural crest cells, limb buds and the central nervous system (Bendall and Abate-Shen, 2000). MSX genes have been shown to act as transcriptional repressors and inhibit differentiation of several cell types (Odelberg et al, 2000; Hu et al, 2001).
If a male swordtail would loose his sword by an injury, he can regrow a sword. As already stated there are different MSX genes. But the MSX-C gene will take care of regenerating a new sword.
Below:
Some female swordtails that have developed a sword. So, these specimens have not changed into a male. And all females show a gravid spot and have all delivered fry.



Above: Female comma swordtail (Xiphophorus signum). Photo courtesy by Albert Palner.
Below: Female Guntheri swordtail (Xiphophorus hellerii guentheri). Photo courtesy by Nigel Hunter.

Below: Female neon swordtail (Xiphophorus hellerii). Photo courtesy by Febri Ariwibowo.

Above: These females are no late males...!
A late male has got nothing to do with the story mentioned before unless it's got to be an aa male of the XX combination. A late male is already an initial male. But because of external factors (e.g. dominance of other fish) they'll grow up large and show the male characteristics very late in comparison to an average male.

Above & below: Two male siblings of the Surinam green guppy strain. The most colored one is of a normal size. The other male is relatively bigger in size than his brother. With most guppies and endlers when a male turns out so big, the colors on the body will fade or remain pale. This is quite normal.

Early males develop when there's hardly a male within a colony. This is a survival system so that the reproduction will be ensured. But also when young specimens are kept in a very warm environment or a too crowded environment, early males will develop.

Above: Two male tuxedo endlers coming from the same colony. The upper male shows the right adult size and coloration. The lower male is a late male which has faded coloration in comparison to the upper one. The only red the bigger shows, is a red stripe on his shoulder.
Polymorphic Melanic Side-Spotting Patterns in Poeciliid Fishes

Above: Representation of the poeciliid pigmentation patterns included in this review. The base pattern lacks macromelanophores, so we interchangeably refer to it as “unspotted.” Melanic side-spotting patterns include any pattern in poeciliid fishes that consists of macromelanophores forming spots that are irregularly distributed on the fish’s flank. Included melanic side-spotting patterns may also extend into fins or be restricted to only a portion of the flank, not pictured here. The black morph is an extreme version of the spotting pattern.
White specimens
In all animal life forms, white and very pale specimens do occur. And by white specimens is meant, lacking pigment or a huge shortage of chromatophores (color cells) in skin, feathers, scales and hair. Many people mix up white appearances with the term "albino". But actually that would be an incorrect statement to make. There are two standards of white appearances in life animals (which includes humans as well). These are albinism and leucism. Albinism can be divided in two standards as well. We have oculocutaneous albinism, which means a specimen which lacks pigment in skin, hair, feathers, scales and eyes (so, total albinism). These are pale or white specimens with red eyes. And we have ocular albinism, which means a specimen which lacks pigment (that could be melanophores or other color cells) in the eyes but it's not lacking pigment in the skin, feathers, scales and hair. So, a specimen with a colored body but with red eyes.
Leucism means a pale or white body with black eyes or other eye colors with the exception of red. In this case the appearance shows a body that's lacking or has a huge shortage of pigment in skin, feathers, scales and hair but with pigment (melanophores or other color cells) in the eyes.

Wild colors / fancy colors
People have their own preferences regarding what the color palette of fish should be. The commercial market focuses mainly on the most colorful fish. And yet there are plenty of people worldwide who prefer wild colors over cultivated colors.
But there are also people who love to mix wild and cultivated colors in their tanks. Most cultivated colors (or also known as fancy colors) were a result of a natural mutation that could have been caused by the environment, available food sources, the presence of predation or a need of adaption in relation to other factors. These color mutations were linebred at some point to create new color varieties of a certain fish species.
It's not debatable why someone should choose for a specimen with wild colors or fancy colors. It's totally a personal preference. But if someone would keep wild and fancy colored fish of the same species in one tank, wild colored specimens will breed with the fancy colored ones. In general, the offspring will be wild colored again (or at least the majority). For wild colors are generally spoken dominant over fancy colors.

Anatomy:
Overhere, an overview of all fins of a male and female ovoviviparous livebearer.


A "majority" of the livebearing species will prefer alkaline water where the pH is between 7.0 and 7.8. In general, the water should be hard or medium hard. But mind that there are livebearer species that do prefer soft water instead of hard water.